霊長類における中心窩視細胞の変動性:視覚的精度を形作る複雑なパターンを探求する。進化と遺伝子が霊長類の視覚における比類のない多様性をどのように促進しているのかを発見する。
- 霊長類における中心窩構造の紹介
- 視細胞研究に関する歴史的視点
- 比較解剖学:霊長類種間の中心窩視細胞
- 視細胞変動性の遺伝的決定因子
- 中心窩構成に影響を与える発達メカニズム
- 視覚的鋭敏度と色覚に対する機能的含意
- 変動性の環境的および進化的要因
- 中心窩視細胞の評価手法
- 臨床的関連性:人間の視覚障害に関する洞察
- 今後の方向性と未解決の質問
- 情報源と参考文献
霊長類における中心窩構造の紹介
中心窩は、霊長類、特に人間の高精細視覚を担う特化した網膜の領域です。この小さな中央のくぼみは、色の詳細な視覚と微細空間解像度に不可欠な視細胞、特にコーン細胞が密集しています。中心窩の構造と細胞構成は、昼行性の霊長類の視覚的要求を支持するように進化しており、彼らが環境内の微妙な色や詳細の違いを検出できるようにしています。しかし、異なる霊長類種間での中心窩視細胞の組織と密度には重要な変動性があり、これは多様な生態的ニッチと視覚的要求への適応を反映しています。
ほとんどの霊長類において、中心窩はコーン視細胞の高い濃度が特徴であり、周辺の網膜ではより一般的で低照度視覚を担当する杆細胞の相対的な欠如があります。中心窩のコーンの密度は、人間では平方ミリメートルあたり最大20万細胞に達し、網膜で最も高い視覚的鋭敏度を持つ領域となっています。しかし、この密度はすべての霊長類種で均一ではありません。たとえば、旧世界のサルや類人猿(人間を含む)は、通常よく発達した中心窩を持ち、はっきりとしたくぼみと高いコーン密度を示しますが、いくつかの新世界のサルは、中心窩の構造があまり明瞭でないか、まれな場合には中心窩がまったく存在しないことがあります。
霊長類間の中心窩視細胞の構成の変動性は、視覚生態における違いと密接に関連しています。食物採取や社会的合図などのタスクにおいて鋭い色覚に大きく依存する種は、より洗練された中心窩の構造を持つ傾向があります。一方、低照度環境に適応した夜行性の霊長類は、しばしば中心窩が減少または欠如しており、杆細胞がより多くを占めています。この多様性は、霊長類の視覚システムを形作る進化的圧力を強調し、中心窩は霊長類の視覚を理解するための重要な解剖学的特徴であることを示しています。
霊長類の中心窩構造に対する研究は、解剖学的研究、in vivoイメージング、遺伝子解析によって進展しており、この網膜特化の開発、機能、および進化的意義に関する洞察を提供しています。国立眼研究所や国立衛生研究所などの組織は、この分野の進行中の研究を支援し、正常な視覚機能や中心窩に影響を与える網膜障害の理解を深めるのに貢献しています。
視細胞研究に関する歴史的視点
霊長類における中心窩視細胞の変動性の研究は、解剖学的技術と視覚システムの概念的理解の両方の進展を反映した豊かな歴史を持っています。19世紀後半と20世紀初頭の初期の調査では、光学顕微鏡を使用して霊長類の網膜の基本的な組織を記述し、特に高精細視覚を担う特化した中央領域である中心窩に注目しました。サンティアゴ・ラモン・イ・カハールのような先駆的な解剖学者は、中心窩におけるコーン視細胞の密集 packing を詳細に描写し、この領域における杆細胞の欠如や、周辺網膜に比べたコーンの独特の伸長と配置を指摘しました。
組織学的な方法が進展するにつれて、研究者たちは異なる霊長類の種間の中心窩コーンの密度と分布を定量化し始めました。これらの研究は重要な種間の変動性を明らかにし、旧世界のサルや類人猿(カタリーネ)が一般的に新世界のサル(プラティリーネ)よりも高い中心窩コーン密度を示すことを示しました。この変動性は、視覚生態や鋭敏視覚に対する行動的依存に関連付けられました。20世紀半ばにおける電子顕微鏡の登場により、より細かい解像度が可能となり、コーンの亜型やそのシナプス接続間の微妙な形態学的差異を特定することができるようになりました。
20世紀後半には、生理学的および心理物理的アプローチが統合され、研究者は解剖学的発見と視覚的鋭敏度や色識別の機能的測定とを関連付けました。コーン視細胞の変動性を基礎付ける遺伝的多型の発見、特にオプシン遺伝子においては、個体および種の差異における分子的基盤を提供しました。これは、霊長類間の三色覚や二色覚の研究において特に注目され、霊長類の視覚の進化理解に深い影響を及ぼします。
近年では、適応光学走査レーザー網膜計測(AOSLO)などの非侵襲的イメージング技術が、人間および非人間霊長類の中心窩コーンモザイクのin vivo可視化を可能にしました。これらの進展は以前の組織学的発見を確認し、コーン密度や配置、まれな視細胞タイプの存在における個人差など、追加の変動の層を明らかにしました。このような研究は、国立眼研究所や国立衛生研究所などの主要な視覚科学組織によってしばしば行われるか支援され、これらの分野の基盤研究の資金提供と普及において重要な役割を果たしています。
全体として、霊長類における視細胞研究の歴史的進展は、技術革新と科学的発見の相互作用を強調しており、中心窩領域の注目すべき変動性と特化の理解を絶えず洗練しています。
比較解剖学:霊長類種間の中心窩視細胞
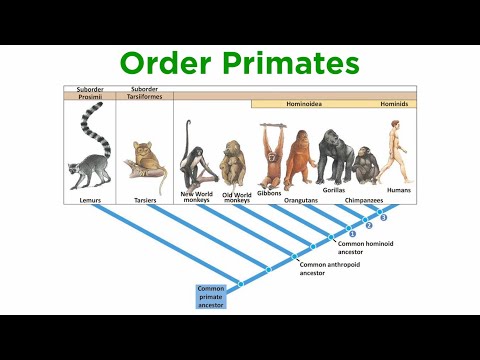
視覚的精度を担う特化した網膜の領域である中心窩は、霊長類の種間で視細胞の構成において顕著な変動性を示します。霊長類において、中心窩はコーン視細胞の密集 packing と杆細胞の相対的欠如によって特徴づけられ、詳細な色認識と空間解像度を最適化しています。しかし、中心窩に存在するコーンの密度、配置、種類は、種によってかなり異なり、多様な生態的ニッチと視覚的要求への適応を反映しています。
人間や他の旧世界のサル(カタリーネ)において、中心窩は高度に発達しており、コーン密度は平方ミリメートルあたり最大20万コーンに達します。これらのコーンは、短波長(S)、中波長(M)、および長波長(L)感受性コーンの3種類に分かれており、三色覚を可能にしています。この配置は微細な視覚識別をサポートし、食物採取や社会的信号の発信などのタスクにおいて有利であると考えられています。これらの種の中心窩のくぼみは深く、よく定義されており、光の散乱を最小化し光学経路を最適化することで視覚的鋭敏度をさらに高めています(国立眼研究所)。
対照的に、新世界のサル(プラティリーネ)は、中心窩の構造や視細胞の構成においてより大きな変動を示します。たとえば、ホエザル(Alouatta)のようないくつかの種は、旧世界の霊長類に似た三色覚を持っていますが、多くの他の種は多型の色覚を示し、X連鎖のオプシン遺伝子の変異によって三色覚を示す女性のサブセットのみが存在します。これらの種における中心窩コーン密度は一般的に低く、中心窩のくぼみはあまり目立たないか、場合によっては欠如していることもあります。この多様性は生息地や食物採取戦略の違いを反映しており、一部の種は無色の手がかりや運動検出により依存していると考えられています(スミソニアン協会)。
レムールやタルシアのような原猿は、真の中心窩を欠き、かわりに中央網膜特化領域であるエリアセントラリスを持ち、コーン密度の中程度の増加を含むものの、霊長類(霊長類)の高い特化には達していません。彼らの視細胞構成はしばしば杆細胞が支配し、限られた色の識別を伴う夜行性または薄明薄暮性のライフスタイルを支えています(アメリカ自然史博物館)。
霊長類間の中心窩視細胞の組織の解剖学的変化は、視覚システムを形作る進化的圧力を強調しています。この変動性は、色覚の要求、空間解像度、光感受信性の間のバランスを反映しており、各種の生態的および行動的文脈に合わせて調整されています。
視細胞変動性の遺伝的決定因子
霊長類の網膜における特化した領域である中心窩は、鋭敏な中心視覚と色の識別を担当する高密度のコーン視細胞によって特徴づけられています。しかし、個々の霊長類、特に人間において、中心窩内の視細胞の数、分布、および種類において顕著な変動性が存在しています。この変動性は、視細胞の発達、分化、および維持を制御する遺伝的決定因子の複雑な相互作用によって影響されます。
中心窩視細胞の変動性に寄与する主要な遺伝的要因の1つは、コーン細胞における光感受性タンパク質をコードするオプシン遺伝子の配列です。人間や他の旧世界の霊長類では、三つの異なるオプシン遺伝子(OPN1LW(長波長)、OPN1MW(中波長)、OPN1SW(短波長)が存在し、三色覚を可能にします。これらの遺伝子の配列、コピー数、および発現の変異は、中心窩内のL、M、およびSコーンの比率や空間的配置の違いにつながる可能性があります。たとえば、X染色体上のOPN1LWとOPN1MW遺伝子間の不均等な再結合イベントは、遺伝子の重複や欠失を引き起こし、コーン比率の個体差や、場合によっては色覚の欠損に寄与することがあります。
オプシン遺伝子の変異に加えて、他の遺伝子座も中心窩の発達や視細胞のパターニングにおいて重要な役割を果たしています。PAX6、CRX、およびNRLなど、網膜の形態形成に関与する遺伝子は、網膜前駆細胞の増殖と分化を調節し、最終的に中心窩におけるコーンの密度や配置に影響します。これらの遺伝子における変異や多型は、特定の遺伝性網膜疾患で見られる構造的な異常や視細胞の分布の変化を引き起こすことがあります。
霊長類の種間比較研究は、遺伝的分岐が中心窩の構造における種間の違いの根底にあることを明らかにしています。たとえば、新世界のサルは、単一のX連鎖のオプシン座の同義体の変異により色覚表現型の範囲を示し、二色覚と三色覚の個体が存在します。対照的に、旧世界の霊長類におけるそれぞれのOPN1LWとOPN1MWの遺伝子を生成する遺伝子重複イベントは、三色覚の安定した基盤とより均一な中心窩コーンモザイクを持つことを確立しました。
ゲノムシーケンシングや単一細胞トランスクリプトミクスの最近の進展は、中心窩視細胞の変動性を統括する遺伝ネットワークについてさらに明らかにしました。これらのアプローチにより、コーン亜型の仕様や空間的組織を微調整するのに寄与する新しい調節要素や遺伝子発現パターンが特定されています。国立眼研究所や国立衛生研究所などの組織によって支援されている進行中の研究は、視覚機能の理解や網膜疾患の治療方法の開発において中心窩の多様性の遺伝的基盤を解明し続けています。
中心窩構成に影響を与える発達メカニズム
中心窩は、霊長類の網膜における特化した領域であり、高精細な視覚にとって重要で、密集したコーン視細胞によって特徴づけられています。しかし、霊長類種間や同種内でも、これらの視細胞の組成や配置には顕著な変動性が存在します。この変動性を駆動する発達メカニズムを理解することは、進化的適応や視覚障害の病因の解明に不可欠です。
網膜の発達中に、中心窩は遺伝的、分子的、環境的要因の複雑な相互作用によって形成されます。中心窩領域の初期の指定は、細胞運命の決定を制御する形態形成因子や転写因子のグラデーションによって調整されます。たとえば、転写因子PAX6の発現は、眼の初期パターニングに重要であり、OTX2やCRXなどの他の因子は視細胞の分化に関与します。これらの分子的手がかりは、網膜前駆細胞の増殖と移動を導き、最終的に中心窩におけるコーンの密度や亜型分布に影響を与えます。
中心窩の発達における重要な側面は、特に長波長(L)および中波長(M)感受性コーンの選択的増加と、中心窩の中心における短波長(S)コーンの相対的な欠如です。このパターンは、内因性の遺伝的プログラムと外因性のシグナル経路の両方を通じて確立されます。たとえば、甲状腺ホルモンのシグナル伝達はオプシン遺伝子の発現を調節し、LコーンとMコーンの比率に影響を与えることが示されています。さらに、前駆細胞の細胞周期からの離脱のタイミングが最終的な視細胞のモザイクに影響を与え、個体間の変動性に寄与する可能性があります。
開発の重要な局面の一つである環境要因、特に開発の重要な期間中の光曝露も、中心窩の構成に役立ちます。非人間の霊長類を対象にした実験的研究は、視覚経験の変化がコーンの密度や配置に影響を与えることを示しており、中心窩の発達における可塑性の程度を示唆しています。さらに、妊娠中の栄養状態や母体の健康状態が、成長因子や栄養素の供給を調整することによって、網膜の発達に間接的に影響を与える可能性があります。
霊長類種間の比較研究は、進化的圧力、たとえば生態的ニッチおよび視覚的要求によって種特有の適応が中心窩構造に導かれることを示しています。たとえば、昼行性の霊長類は、夜行性の種に比べて通常は高い中心窩コーン密度を示し、彼らの環境における色覚と視覚的鋭敏度の重要性を反映しています。これらの違いは、遺伝的遺産と適応反応が中心窩視細胞の変動性を形成する影響を強調しています。
国立眼研究所や国立衛生研究所などの組織が支援する進行中の研究は、中心窩の構成を根底から成り立たせる複雑な発達メカニズムを解明し続けています。これらの研究からの洞察は、霊長類の視覚の理解を向上させるだけでなく、中心窩に影響を与える網膜疾患の診断や治療戦略にも役立ちます。
視覚的鋭敏度と色覚に対する機能的含意
中心窩は、霊長類の網膜における特化した領域であり、コーン視細胞が密集しており、高解像度の視覚と色の識別にとって重要です。霊長類種間、さらには個々の中心窩視細胞の密度、分布、タイプの変動性は、視覚的鋭敏度と色覚の両方に対して大きな機能的含意を持っています。
視覚的鋭敏度とは、微細な空間的詳細を解像する能力を指し、中心窩におけるコーン視細胞の密度によって直接的に影響を受けます。人間や他の旧世界の霊長類では、中心窩のコーン密度は平方ミリメートルあたり最大20万コーンに達し、動物界でも最高レベルの空間解像度を支えています。しかし、この密度はすべての霊長類で均一ではありません。たとえば、新世界のサルはしばしば中心窩コーン密度が低く、これは一般的に低い視覚的鋭敏度と相関しています。同一種でも、中心窩のコーンの詰まりの固有の違いが視覚性能に測定可能な違いをもたらす可能性があります。これらの変動は、遺伝的要因、発達的要因、さらには生態的ニッチに適応した進化的適応によって生じると考えられています。
霊長類の色覚も中心窩視細胞の変動性の影響を受けます。ほとんどの霊長類は、短波長(S)、中波長(M)、および長波長(L)感受性コーンの3種類のコーン視細胞を持ち、三色覚を実現しています。これらのコーンの相対的な比率と中心窩における空間的配置は、色識別能力に影響を及ぼします。たとえば、人間では、LコーンとMコーンの比率は個々の間で広く異なるが、ほとんどが強力な色覚を保っており、神経機構が視細胞の変動性を補償していることを示唆しています。対照的に、いくつかの新世界の霊長類は多型の色覚を示し、X連鎖のオプシン遺伝子の変異によりまれに三色覚を持つ女性のサブセットが存在し、他の個体は二色覚となります。この遺伝的多様性は、個体間の色の知覚や、食物採取のような生態的行動に顕著な差異をもたらします。
中心窩視細胞の変動性には、臨床的な文脈にも影響を及ぼします。コーン密度や配置の変化は、色覚の欠損や視覚の距離の低下と関連している可能性があり、進化生物学と医学の両方でこれらの違いを理解する重要性が強調されます。継続中の研究は、国立眼研究所や国立衛生研究所などの組織によって支援され、霊長類の視覚における中心窩視細胞の変動性やその影響を解明し続けています。
変動性の環境的および進化的要因
霊長類における中心窩視細胞の変動性は、環境的および進化的要因の複雑な相互作用によって形作られています。高精度視覚を担う特化した網膜領域である中心窩は、視細胞の密度、配置、構成において種間および個体内で顕著な違いを示します。これらの違いはランダムではなく、生態的ニッチ、視覚的要求、および進化の歴史に密接に結びついています。
中心窩の変動性の主要な環境要因の1つは生息地のタイプです。密生した森林に生息する霊長類、特に多くの新世界のサルは、しばしば低照度条件や複雑な視覚環境に直面しています。これらの環境では、感度を向上させるために杆細胞の比率が高まるか、またはまだ配光条件による色識別に最適な特定のコーンの配置が選抜される可能性があります。対照的に、開けた生息地(サバンナなど)に住む霊長類はより明るく均一な照明に晒されており、高いコーン密度と密接に詰まった中心窩の進化を導くことがあります。これは捕食者検出や小さくカラフルな果物や昆虫を食べるための急速な視覚的タスクをサポートするためです。
食事に特化することも、中心窩視細胞の構成に選択圧を加えます。色覚で熟した果物を特定することに重きを置く果物食性霊長類は、長波長に感受性の高いコーン視細胞の多様性と密度が高いことがよくあります。この適応は、彼らが環境内の微妙な色の違いを識別する能力を向上させます。一方、主に葉を食べるfolivorous種は、詳細な色覚がそれほど必要ではないため、異なる中心窩視細胞のプロファイルを持つことがあります。
進化的系統も変動性に寄与します。旧世界のサルや類人猿(カタリーネ)は、通常、詳細な視覚情報に依存するため、十分に発達した中心窩を持ち、高いコーン密度を有します。対照的に、多くの原猿や一部の新世界のサル(プラティリーネ)は、中心窩の特化が少なく、進化的圧力や先祖の視覚的要求を反映しています。遺伝子研究は、オプシンタンパク質に影響を及ぼす遺伝子の重複や変異が、霊長類の色覚の多様化と中心窩の構造に重要な役割を果たしてきたことを明らかにしました。
最後に、顔認識や複雑な社会的信号の必要性など、社会的および行動的要因が中心窩視細胞の配置に影響を与える可能性もあります。複雑な社会システムを持つ種は、微妙な顔の手がかりを解釈するために急速な視覚を必要とし、高い中心窩コーン密度の進化を促進する可能性があります。
これらの環境的および進化的要因は、霊長類における中心窩視細胞の変動性の適応的意義を強調しており、生態的要求と系統的制約との間の動的なバランスを反映しています。国立衛生研究所やNature Publishing Groupなどの組織による進行中の研究は、この顕著な多様性の根底にある遺伝的および発達的メカニズムを解き明かし続けています。
中心窩視細胞の評価手法
霊長類における中心窩視細胞の変動性を評価するには、高度なイメージング、組織学的、分子技術の組み合わせが必要です。これらの手法は、高精度視覚を担う特化した網膜領域である中心窩内の視細胞、主にコーンの微細構造と分布を捉えることを目的としています。手法の選択は、研究の目的、調査する種、およびin vivoまたはex vivoの分析が必要かどうかに依存します。
最も広く使用される非侵襲的技術の1つは、適応光学走査式光網膜計測(AOSLO)です。この技術は、眼の光学的なひずみを修正し、生活している霊長類の個々の視細胞を高解像度でイメージングすることを可能にします。AOSLOにより、研究者は中心窩内のコーンの空間的配置と密度をマッピングし、時間の経過に伴う変化を追跡し、個体間の変動性を比較することができます。この技術は、霊長類種間や同一種の個体間でのコーンの配置や分布における微細な違いを明らかにする上で重要な役割を果たしています。
光干渉断層撮影(OCT)、特にスペクトル領域およびスイープソース変法は、マイクロメートル解像度で網膜の断面画像を提供します。OCTはAOSLOほど個々の視細胞を明確に解決できませんが、中心窩の厚さ、層の完全性、および中心窩のくぼみ全体の構造を測定するのに非常に役立ちます。これらの構造的パラメータは、視細胞の密度や組織と相関し、中心窩の変動性へ間接的かつ補完的な洞察を提供します。
Ex vivoの研究では、組織学的分析が金標準となっています。網膜組織は固定され、断面を作成し、光学顕微鏡または電子顕微鏡の下で視細胞を視覚化するために染色されます。免疫組織化学は、特定のオプシンタンパク質を標的にすることによって、コーン亜型(例えば、S、M、Lコーン)間をさらに区別することができます。このアプローチは、視細胞の正確な数と空間的マッピングを提供しますが、死後のサンプルに制限され、組織処理のアーチファクトに影響される場合があります。
分子技術(in situ ハイブリダイゼーションや単一細胞RNAシーケンシングなど)が、中心窩視細胞の遺伝的およびトランスクリプトミクスの多様性を評価するためにますます使用されています。これらの方法は、コーン間の機能的変動性を基づく遺伝子発現の微妙な違いを特定することができ、霊長類における中心窩の特化の分子的基盤をより深く理解するのに寄与します。
これらの手法、すなわち高解像度のin vivo イメージングから詳細な分子的プロファイリングまでが集約されることで、中心窩視細胞の変動性の包括的評価が可能になります。これらは霊長類の視覚の理解を深めるために不可欠であり、眼科および神経科学における応用研究を情報化するのに役立ちます。これらの手法を支援し、標準化する主要な組織には、国立眼研究所や視覚と眼科学の研究会が含まれており、視覚科学研究と普及において重要な役割を果たしています。
臨床的関連性:人間の視覚障害に関する洞察
中心窩は、霊長類の網膜における特化した領域であり、密集したコーン視細胞が存在し、高精度視覚にとって重要です。霊長類、特に人間における中心窩の視細胞密度、分布、亜型の変動性には、視覚障害の理解と診断において重大な臨床的含意があります。中心窩の独自の構造は、高濃度のコーンと杆細胞の欠如によって特徴づけられており、それが色の識別と物体の空間的解像を支える役割を果たしています。しかし、中心窩視細胞の配置における個体差は、さまざまな網膜疾患に対する感受性やその発症に影響を与える可能性があります。
中心窩視細胞の変動性に関連する最も臨床的に重要な側面は、遺伝性の網膜疾患との関連です。たとえば、色覚異常、コーンジストロフィー、および黄斑変性症などの状態は、中心窩でのコーン視細胞の選択的な喪失または機能不全を含むことがよくあります。視細胞の喪失の程度や影響を受ける特定の亜型(L、M、またはSコーン)の選択は、色盲から重度の中心視力の喪失まで、視覚欠損のスペクトルをもたらす可能性があります。健康な個体間における中心窩コーン密度や配置の自然な変動性を理解することは、病的変化を正常な解剖学的差異から区別する上で重要な基準を提供します。
適応光学走査レーザー網膜計測などの高解像度網膜イメージングの最近の進展により、臨床医や研究者はin vivoで中心窩視細胞モザイクを視覚化し定量化することが可能になりました。これらの技術は、正常な視覚を持つ個体の間でさえ、中心窩のコーン密度や配置の規則性にかなりの変動があることを明らかにしました。このような発見は、網膜疾患の診断およびモニタリングにおいて個別のアプローチの重要性を強調しており、集団平均からの逸脱が必ずしも病理を示すものではないことを示しています。
人間と類似の中心窩の構造と機能を持つ非人間の霊長類における比較研究は、視細胞の変動性に関連する遺伝的および発達的要因をさらに明確にしました。これらのモデルは、中心窩の疾患を対象とした遺伝子および細胞ベースの治療の前臨床テストにおいて非常に価値があります。さらに、種間の違いを理解することは、動物モデルから人間の臨床実践への発見の翻訳を助けるうえで重要です。
最終的に、中心窩視細胞の変動性に関する洞察は、臨床的イメージングの解釈、診断基準の改善、および中心窩および黄斑疾患に向けたターゲット治療の開発を向上させる能力を高めます。国立眼研究所や世界保健機関などの組織が支援する進行中の研究は、健康と疾患における中心窩の役割に関する知識をさらに広げており、視覚障害を持つ患者の予後を改善する道を開いています。
今後の方向性と未解決の質問
霊長類の中心窩の構造と機能に関する理解が大きく進展したにもかかわらず、この特化した網膜領域における視細胞の変動性に関して多くの問いが残っています。今後の研究は、これらの変動性の根底にあるメカニズムとその機能的結果の両方を解決することを目指しており、視覚科学、進化生物学、臨床眼科学において重要な意味を持っています。
1つの主要な関心のある分野は、中心窩視細胞の変動性の遺伝的および発達的基盤です。コーンの密度や分布が個体間および霊長類種間でかなり異なることが確立されていますが、これらの違いを駆動する正確な遺伝的要因や分子的経路は完全には解明されていません。単一細胞トランスクリプトミクスやゲノム編集技術の進展により、特定の遺伝子や調節要素が中心窩の構造や視細胞亜型の仕様に寄与する様子を解明することが可能になるでしょう。
もう1つの重要な質問は、中心窩の変動性の適応的意義に関するものです。霊長類の系統間の比較研究は、生態的要因(食事や生息地など)が中心窩の構造や視細胞の組成の進化に影響を与える可能性を示唆しています。しかし、環境圧力、視覚的要求、および視細胞の構成間の直接の関連はまだ明らかではありません。縦の調査や種間研究(非侵襲的イメージングモダリティを利用する可能性)により、自然選択によって中心窩の特性がどう形成され、異なる生態的文脈での視覚的性能にどう寄与するかを明らかにすることができるでしょう。
高解像度の網膜イメージングにおける技術的進歩(適応光学走査レーザー網膜計測など)は、今後の調査において重要な役割を果たすと期待されています。これらのツールは、個々の視細胞のin vivo可視化と定量化を可能にし、研究者が前例のない空間的スケールで変動性をマッピングすることを可能にします。構造の変動性と知覚結果を結びつけるためには、機能的評価(心理物理テストや電気生理学的記録など)とのデータ統合が重要です。
また、中心窩視細胞の変動性が網膜疾患に与える影響に関して未解決の質問も残っています。個体間の中心窩の構造的違いが、加齢黄斑変性症や遺伝性網膜ジストロフィーのような状態の感受性や進行にどう影響を与えるのかはまだ明らかではありません。大規模で縦の調査が、人間と非人間の霊長類の両方でこれらの関係を探求し、診断や治療に向けての個別化アプローチを情報化するために必要です。
最後に、視覚科学者、遺伝学者、臨床医の間での共同作業は、国立眼研究所や国立衛生研究所などの組織によって支援され、これらの複雑な質問に取り組む上で重要です。研究が中心窩視細胞の変動性の複雑さを解明し続ける中で、基本的な科学と臨床ケアの両方を進展させる新しい洞察が出現する可能性があります。
情報源と参考文献